
Categoría: Investigación Científica y Tecnológica
Artículo Original
Reconstrucción de la filogenia de las especies de líquenes colombianas del género sticta: efecto de la actualización de los programas de análisis molecular evolutivo
Alejandro Noguera Gómez 1, Alejandro Yepes Bolívar 2, Mario A. Gómez Ortegón2, Tomás Uribe Carrasco 2 y Mauricio Pulido Jiménez 3
1. Joven Investigador CEBM. 2. Estudiantes 11º grado, Gimnasio Campestre. 3. Director CEBM, Gimnasio Campestre
Correspondencia para el autor: mpulido@campestre.edu.co
Recibido: 21 de marzo de 2023
Aceptado: 5 de mayo de 2023

RESUMEN
ABSTRACT
Utilizando un listado de secuencias ITS y programas para análisis de evolución molecular actualizados se reconstruyó el árbol filogenético de las especies colombianas de líquenes pertenecientes al género Sticta propuesto en trabajos anteriores. El número y la disposición de los agrupamientos (clados) que muestra el nuevo árbol mostraron variaciones respecto del elaborado en investigaciones previas. Las accesiones que conformaron los nuevos clados quedaron claramente separadas de aquellos a los que pertenecían inicialmente, hecho cuyo grado de fiabilidad está respaldado por los valores de soporte estadístico que mostraron los análisis.
Palabras clave: liquen, género Sticta, secuencia ITS, árbol filogenético, clado.
Using an index of ITS sequences and updated programs for molecular evolution analysis, the phylogenetic tree of Colombian lichen species belonging to the genus Sticta proposed in previous works was reconstructed. The number and arrangement of the groupings (clades) shown in the new tree showed variations with respect to the one elaborated in previous research. The accessions that formed the new clades were clearly separated from those to which they initially belonged, a fact whose degree of reliability is supported by the statistical support values shown by the analyses.
Key words: lichen, genus Sticta, ITS sequence, phylogenetic tree, clade.
Desde el punto de vista botánico, los líquenes son el resultado de una asociación simbiótica entre dos o más comunidades fotoautótrofas de algas y cianobacterias, llamadas fotobiontes, y un hongo, llamado micobionte

INTRODUCCÍON
En la corteza de los árboles, cubriendo superficies rocosas o incluso en estructuras artificiales, habitan organismos únicos para la ciencia y útiles para diversos propósitos pero que suelen pasar inadvertidos para la mayoría de los humanos: los líquenes. Desde el punto de vista botánico, los líquenes son el resultado de una asociación simbiótica entre dos o más comunidades fotoautótrofas de algas y cianobacterias, llamadas fotobiontes, y un hongo, llamado micobionte (Hawksworth, 1989). Este tipo de organización simbiótica presupone que el micobionte es responsable de la reproducción del organismo mediante esporas, mientras que los fotobiontes son responsables de la captación de energía y la producción de alimento mediante fotosíntesis. La función fotosintética puede ser realizada por dos tipos de organismos: las clorofitas (algas verdes) o las cianobacterias, organismos procarióticos capaces de realizar fotosíntesis oxigénica (Spribille, et al., 2022). Las clorofitas son un grupo de organismos eucarióticos que contienen clorofila como principal pigmento fotosintético, es decir, que utilizan la luz solar para producir energía química. Por otro lado, las cianobacterias contienen pigmentos llamados ficobilinas que les permite absorber radiación lumínica de diferentes longitudes de onda. De este modo, pueden realizar la fotosíntesis en entornos con poca luz y en aguas profundas (Mirande y Tracanna, 2007). A su vez, organismos pertenecientes a tres divisiones del reino Fungi suelen participar en la asociación: ascomicetos, basidiomicetos o ficomicetos (Alexopoulos y Mims, 1985). En función de todo lo anterior, la taxonomía de los líquenes resulta compleja dado que no son protistas pero tampoco pertenecen al reino de los hongos.
Los líquenes son considerados bio-indicadores puesto que responden a diferentes grados de alteración de los elementos que hacen parte del entorno en el que habitan, como el aire, los suelos y el agua, entre otras, a través de cambios morfológicos o funcionales que pueden ser identificados, como cambios de coloración o producción de esporas para la reproducción (García y Rubiano, 1984). Un análisis de la presencia o ausencia de ciertas especies liquénicas en un entorno particular puede ofrecer información sobre las condiciones del ecosistema en el que se encuentran. Por ejemplo, la ausencia de una especie liquénica puede ser indicio de alteraciones o problemas en el ecosistema; esto se debe a que algunos líquenes no logran adaptarse a condiciones desfavorables, como la presencia de aguas contaminadas con aceites y demás compuestos orgánicos o químicos. Su comportamiento ante tales condiciones difiere del natural en aspectos tanto fisiológicos como poblacionales, lo cual provoca la prevalencia de ciertas especies y subespecies con mayor adaptabilidad a condiciones adversas (Acevedo y Charry, 2018). La presencia de líquenes, analizada desde la perspectiva de su diversidad (tanto morfológica como genética), permite conocer las condiciones de un ecosistema.
Los espaciadores transcritos internos (ITS, por sus siglas en inglés) son pequeñas secuencias genómicas presentes en el micobionte, específicamente en el ADN ribosómico, y tienen alrededor de 250 pb de longitud. Las secuencias nucleotídicas de estos segmentos del genoma fúngico muestran una gran variabilidad, razón por la cual son de gran utilidad en estudios genéticos y evolutivos. El uso de estos segmentos de DNA es una de las herramientas más frecuentemente utilizadas para determinar los niveles de variabilidad genética entre organismos que hacen parte de una población.
En la actualidad, los estudios de variabilidad genética son importantes en la medida en que aportan información esencial a otras áreas de la biología, como la taxonomía. El análisis taxonómico de los seres vivos consiste en el estudio detallado de los caracteres morfológicos que permiten distinguir a una especie de otra; mediante este tipo de análisis, junto con el uso de la información de secuencia de cierto tipo de genes, es posible formular hipótesis sobre las relaciones de parentesco que existen entre las especies. A través del estudio de los resultados de estos análisis se pueden identificar los vínculos evolutivos entre los organismos. La representación gráfica de las relaciones de parentesco entre individuos de una población se conoce como cladograma. Los individuos más similares entre sí están agrupados en arreglos de disposición ramificada denominados clados, de los cuales se deriva el nombre del diagrama. Estas representaciones visuales no son nada diferente a la manifestación de las distancias genéticas que hay entre los individuos de una población; la idea de distancia genética se debe entender como la proporción de cambios que muestra una secuencia genética en una posición particular respecto a otra (Lemey, et al., 2009).
En Colombia, los conocimientos biológicos básicos de los recursos naturales son escasos, especialmente en el caso de los líquenes. Dada la complejidad de la taxonomía liquénica, se considera que una clasificación basada en caracteres morfológicos no es suficiente para realizar análisis evolutivos. Este estudio pretende generar el conocimiento necesario para dilucidar las relaciones evolutivas entre las especies liquénicas del género Sticta existentes en el territorio colombiano. Para este fin se analizaron las secuencias ITS de los ejemplares estudiados por Moncada et al., (2013) y que se encuentran registradas en la base de datos Nucleotide. Adicionalmente se hizo uso de la información correspondiente a algunas accesiones externas que se emplearon como grupo de control.
El propósito del presente trabajo de investigación es analizar computacionalmente las relaciones filogenéticas existentes entre las especies del género Sticta de origen colombiano y establecer un contraste con los resultados reportados por Moncada et al., (2013).

MATERIALES Y MÉTODOS
Listado de secuencias ITS de especies del género Sticta
Preparación de las secuencias para el análisis de filogenia
Alineamiento de las secuencias nucleotídicas
Construcción del árbol de filogenia
Comparación con estudios previos
Las secuencias ITS correspondientes a las especies analizadas se obtuvieron de la base de datos Nucleotide de NCBI. Para este estudio se consideraron las secuencias pertenecientes a los especímenes colombianos reportados por Moncada et al., (2013) y como grupo externo se utilizaron secuencias pertenecientes a especímenes de los Estados Unidos.
Todo trabajo de filogenia requiere que las secuencias que se pretende estudiar se introduzcan al programa de análisis de datos organizadas de manera particular. Las 288 secuencias ITS seleccionadas fueron descargadas de Nucleotide y organizadas en un archivo matriz con formato fasta (Becedas, 2021). Considerando que las secuencias seleccionadas tenían longitudes variables, fue necesario hacer un proceso de edición manual con el fin de igualarlas.
Con el fin de alinear las secuencias seleccionadas para el estudio se empleó el algoritmo MUSCLE, que está disponible dentro del programa Mega X.
El archivo con las secuencias alineadas fue procesado en el programa Mega X mediante el uso de los algoritmos de Máxima Verosimilitud y UPGMA, los cuales se configuraron para realizar 1000 y 500 iteraciones, respectivamente. El procesamiento de datos produjo árboles de filogenia que muestran los valores de soporte de cada muestra.
El árbol consenso, que se entiende como el diagrama final producido por el programa y que fue generado con el algoritmo UPGMA fue seleccionado para los análisis subsiguientes, dado que este presentó valores de soporte más altos que el de máxima verosimilitud, indicando una fidelidad estadística mayor. Este se contrastó con el presentado por Moncada et al., (2013) para identificar los clados coincidentes en ambos diagramas y las diferencias entre ellos.
RESULTADOS Y DISCUSIÓN

Para el análisis que se presenta a continuación debe tenerse en cuenta que el árbol filogenético 1 (Aquí va el hipervínculo que lleva a la imagen interactiva del diagrama en formato PDF) corresponde al diagrama que se obtuvo del procesamiento de los datos con el algoritmo UPGMA (500 iteraciones), mientras que el árbol filogenético 2 (Aquí va el hipervínculo que lleva a la imagen interactiva del diagrama en formato PDF) corresponde al diagrama que se obtuvo del procesamiento de los datos con el algoritmo de Máxima Verosimilitud (1000 iteraciones). Este análisis se hizo en sentido ascendente, es decir, de abajo hacia arriba, debido a que las accesiones foráneas de control, en ambos diagramas, se encuentran en la parte inferior del árbol filogenético.
Al observar el árbol consenso de los algoritmos UPGMA (Aquí va el hipervínculo que lleva a la imagen interactiva del diagrama en formato PDF) y de Máxima Verosimilitud (MV) (Aquí va el hipervínculo que lleva a la imagen interactiva del diagrama en formato PDF), se identificó una clara división entre las accesiones colombianas del género Sticta y las foráneas pertenecientes al género Usnea. Estas se utilizaron como grupos externos para verificar el buen funcionamiento del programa Mega X. Por tratarse de un género diferente y provenir de localizaciones geográficas distintas, estas especies están claramente diferenciadas de las especies locales desde el punto de vista morfológico y genético (Suárez y Lücking, 2013), hecho que implica que el análisis las agrupa en un mismo clado.
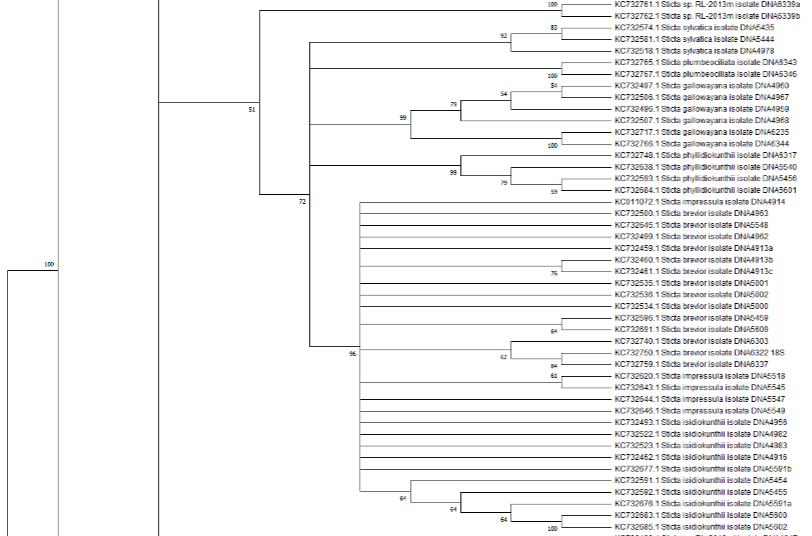
Arriba del clado de control hay diez accesiones que aún no han sido determinadas y que están identificadas con el código sp.RL-2013K y sp.RL-2013T. Dada la falta de información, no está claro si se trata de accesiones de la misma especie colectadas en diferentes localidades geográficas o si son accesiones pertenecientes a especies diferentes que comparten una serie de similitudes genéticas (figura 2).
La siguiente agrupación del género Sticta está formada por seis accesiones: tres ejemplares de la especie S. hirta y tres ejemplares de la especie S. minutula. En este clado, una accesión de la especie S. hirta muestra una mayor cercanía con las accesiones de S. minutula que con los de su propia especie. Las anteriores accesiones forman parte de un clado más grande denominado por Moncada et al., (2013) como Sticta fuliginosa, el mayor de este género, puesto que incluye 20 especies. Según Moncada et al., (2013) las accesiones de Sticta gyalocarpa, las cuales están ubicadas junto a la agrupación de ejemplares no identificados (RL-2013K y sp.RL-2013T), deberían formar parte de esta primera agrupación. Sin embargo, en nuestro árbol encontramos que S. gyalocarpa se aleja de ejemplares que para Moncada et al., (2013) son genéticamente similares, como S. minutula, S. hirta y S. arbuscula.
Asimismo, aislamientos de las especies S. arbusculomentosa y S. tunjensis hacen parte del clado S. fuliginosa. No obstante, estos ejemplares son similares a dos aislamientos de especie aún indeterminada, al igual que una accesión de S. fuliginosa relacionada con S. tunjensis y S. leucoblepharis. Genéticamente similar a S. tomentosa y S. leucoblepharis, se encuentra una accesión de una especie no determinada que hace parte del mismo clado. Sumado a lo anterior, en el mismo clado se encuentran las especies S. papillata, S. dilatata y S. liumbischiana; la última de estas, si bien se encuentra registrada en nuestro árbol filogenético, no aparece registrada por Moncada et al., (2013) como parte de la agrupación. Después del clado de S. fuliginosa hay una agrupación de especies aún no determinadas que está conformado por doce accesiones, registradas en el nuevo árbol construido.
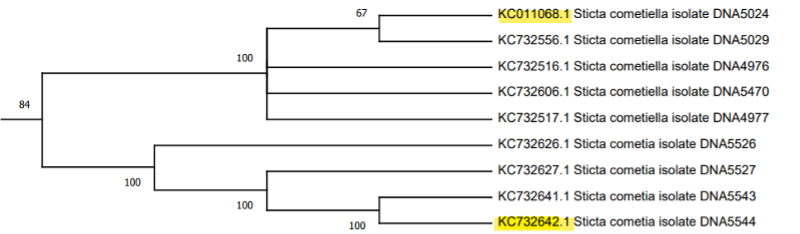
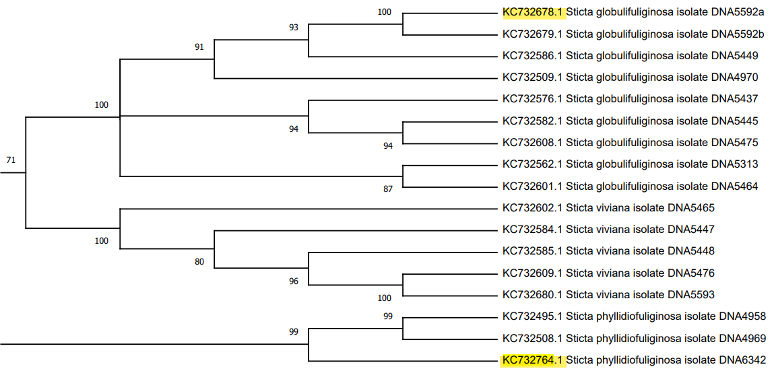
El siguiente clado, descrito por Moncada et al., (2013) como S. kunthii, incluye a las especies S. isidiokuntii, S. impressula, S. brevior, S. phylidiokunthii, S. gallowayana, S. plumbeociliata y S. sylvatica. Enseguida está el clado S. cometia, que incluye cuatro ejemplares de S. cometia y cinco ejemplares de S. cometiella, seguido por un clado de nueve accesiones pertenecientes a una especie no determinada. Sin embargo, en este clado Moncada et al., (2013) describe una especie adicional denominada S. duplolimbata. Por otro lado, el trabajo anteriormente citado describe el clado Sticta viviana, que tiene accesiones de las especies S. phyllidofuliginosa, S. viviana y S. globulifuliginosa, cada una con tres, cinco y ocho accesiones, respectivamente.
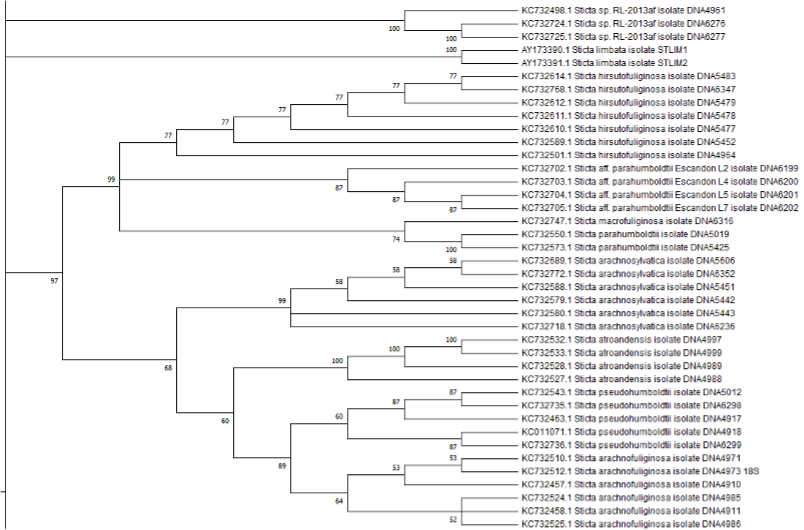
Genéticamente cercano al grupo anterior está el clado S. humboldtii, compuesto por once accesiones diferentes: S. limbata, S. atroandensis, S. parahumboldtii, S. macrofuliginosa, S. pseudohumboldtii, S. hirsutofuliginosa, S. arachnofuliginosa y S. arachnosylvatica, además de tres accesiones no determinadas. Por otro lado, Moncada et al., (2013) describe las especies S. pseudosylvatica, S. aff. ambavillaria y S. humboldtii. Asimismo, evidencia una relación genética cercana (como especies hermanas) entre S. pseudohumboldtii y S. arachnofuliginosa. Sin embargo, a pesar de ser monofiléticas las especies tienen características morfológicas distintas (Moncada et al., 2013); de hecho, “S. pseudohumboldtii tiene una superficie de lóbulos uniformes con tomento denso y sólo produce apotecios, y su superficie es de color claro, mientras que S. arachnofuliginosa tiene una superficie fuertemente faveolada con tomento desarrollado principalmente en las depresiones y produce isidios, y su superficie es de color gris-marrón oscuro.” (Moncada et al., 2013).
El siguiente clado, S. andensis, está conformado por una accesión de la especie S. andensis y cuatro accesiones no determinadas. Moncada et al., (2013) describen el clado S. macrothallina, constituido por tres ejemplares de esta especie. El siguiente clado es denominado en la investigación anteriormente referida como S. canariensis y se compone de accesiones pertenecientes a las especies S. jaguirreana, S. pseudolobaria, S. lobarioides, S. macrofuliginosa y S. neopulmonarioides, además de cuatro accesiones no determinadas. Las especies S. canariensis, S. fragilinata, S. carolinensis y S. ocaniensis son descritas por Moncada et al.,(2013) como parte de esta misma agrupación, mientras que en el árbol 1 no se encuentran. Adicionalmente, se observa el clado S. weigelii, constituido por siete accesiones pertenecientes a las especies S. weigelii y S. tatamana. Sin embargo, en oposición a lo observado en el árbol 1, Moncada et al., (2013) incluye a la especie S. laminobeauvoisii.


El siguiente clado se denomina S. subscrobiculata y contiene ocho accesiones no determinadas, además de S. subscrobiculata y S. laminobeauvoisii, la cual es descrita por Moncada et al., (2013) como parte de un clado ubicado anteriormente, que, sin embargo, no se encuentra en los diagramas obtenidos. El siguiente clado, morfológica y genéticamente relacionado con el anterior, se denomina S. pulmonarioides y reúne accesiones de S. pulmonarioides y especies afines. Adicionalmente, uno de los subclados de S. pulmonarioides contiene una serie de accesiones no determinadas. Existe también un clado con dos accesiones de la especie S. aff. Subscrobiculata.
A continuación, Moncada et al., (2013) describe 11 clados compuestos por 23 especies, que pertenecen a los morfotipos Sticta weigelii y S. carolinensis. Entre estos se encuentran S. andina, S. beauvoisii, S. paramuna, S. phyllidata y especies afines a la anterior. A diferencia de lo obtenido en el árbol 1, Moncada et al., (2013) describe también como parte del morfotipo a las especies S. colombiana, S. dioica, S. fragilinata, S. hypoglabra, S. isidiokunthii, S. laminobeauvoisii, S. laselvae y S. phyllidiokunthii.
Después de la comparación entre los resultados de Moncada et al., (2013) y el árbol filogenético consenso producido en este estudio, se expondrán las generalizaciones y deducciones a partir del contraste entre nuestros resultados y los anteriormente referenciados. En primer lugar, vale la pena recordar que se tomó la decisión de realizar el análisis de datos considerando los resultados representados en el árbol filogenético 1, el cual fue elaborado con el programa Mega X y utilizando el algoritmo UPGMA (500 repeticiones). En el árbol de máxima verosimilitud (MV) de 1000 repeticiones los valores de soporte de cada nodo eran menores a 50, mientras que el de 500 repeticiones presentó cifras mayores a 50, lo que implica que este último muestra un soporte estadístico más robusto.
Los árboles filogenéticos aquí presentados fueron el resultado de programas de procesamiento estadístico actualizados y optimizados que han mejorado su efectividad desde el artículo de Moncada et al., (2013). Prueba de ello es la comparación entre los valores de soporte de los árboles filogenéticos 1, 2 y el de Moncada et al., (2013), pues en el último caso sus valores de soporte se expresan en dos rangos: uno que va entre el 70% y el 90% y otro que va entre 90% y 100% (Suárez y Lücking, 2013). La gran mayoría de los valores de soporte reportados por Moncada et al., (2013) están dentro de un rango que oscila entre el 70% y el 90%, mientras que en el caso de los árboles que se presentan en este estudio están en promedio entre 80% y 100%.
Habiendo comparado los árboles del estudio de referencia y los generados por esta investigación, es posible establecer similitudes entre los árboles 1 y 2. Ambos comparten un clado constituido por ejemplares no determinados, los cuales tienen, en el caso del árbol 2, las letras u y aj y en el caso del árbol 1, k como terminales del código. Este clado es denominado por Moncada et al., (2013) como S. marilandia y la organización del clado es igual en ambos árboles. El siguiente agrupamiento en el árbol filogenético 2 es denominado por Moncada et al., (2013) como S. pulmonarioides, el cual, en este arbol está ubicado al comienzo del diagrama en un orden ascendente, a diferencia del árbol filogenético 1, en donde esta posición la lleva el clado S. fuliginosa.
A continuación, en el árbol filogenético 2 se observa un clado independiente compuesto por dos accesiones de especies cercanas o afines a S. subscrobiculata. Enseguida está el clado S. beauvoisii, constituido por tres accesiones de la misma especie y tres accesiones de S. lobulata. El agrupamiento que se encuentra junto al anterior fue denominado por Moncada et al., (2013) como S. rhizinata y muestra seis accesiones de la misma especie y una especie afín a S. cordillerana. Posteriormente, hay un clado integrado por seis accesiones no determinadas, que por su ubicación indican similitud con un clado denominado por Moncada et al., (2013) como S. hypoglabra. El clado denominado S. weigelii se compone por dos accesiones de la especie S. weigelii, un ejemplar de la especie S. tatamana y cinco accesiones de una especie no determinada.
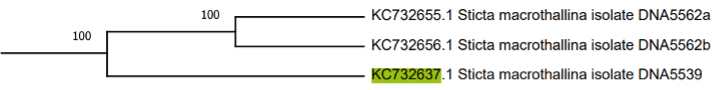
El siguiente clado, S. colombiana, es el de mayor tamaño y se compone por veinte subclados constituidos por las especies S. andina, S. paramuna, aff. phylidiata y accesiones no determinadas. A continuación, se encuentra el clado S. scabrosa, conformado por un sinnúmero de accesiones de especies no determinadas; se denomina así pues Moncada et al., (2013) encuentran similitudes entre los ejemplares y el morfotipo de S. scabrosa. El clado adyacente es denominado por Moncada et al., (2013) como S. canariensis y se compone de las especies S. neopulmonarioides, S. macrofuliginosa, S. pseudolobaria, S. jaguirreana, S. lobarioides y ejemplares no determinados. Posteriormente se encuentra S. fuliginosa, un clado de gran tamaño que se compone de dos subclados: S. tomentosa y S. gyalocarpa, junto con una accesión que pertenece a la especie S. tunjensis. Cada uno de los subclados antes mencionados presenta un número mayor a quince accesiones pertenecientes a especies afines a las que conforman este mismo clado en el árbol filogenético 1. Luego se encuentra el clado que agrupa a las especies de control, seguido por un clado conformado únicamente por tres accesiones pertenecientes a S. macrothallina (Suárez y Lücking, 2013).
El clado S. humboldtii está conformado por nueve subclados de las especies S. hirsutofuliginosa, S. aff. parahumboldtii, S. macrofuliginosa, S. parahumboldtii, S. atroandensis, S. arachnosylvatica, S. pseudohumboldtii, S. arachnofuliginosa y S. macrothallina. El siguiente clado, S. andensis se compone por cuatro accesiones de una especie no determinada junto con un ejemplar de la especie S. andensis. Luego se encuentra S. viviana, un clado compuesto por cuatro subclados de las especies S. viviana y S. globulifuliginosa (Suárez y Lücking, 2013). Posteriormente, se ubica el clado S. albocyphellata que reúne a nueve accesiones de especies indeterminadas y que, en el árbol de referencia, tiene una ubicación similar. Junto a la agrupación anterior, se encuentra el clado S. cometia que está constituido por dos subclados, uno que corresponde a la especie S. cometia y el otro a la especie S. cometiella. El último clado y el de mayor tamaño es S. kunthii, que incluye a seis subclados de las especies S. brevior, S. impressula, S. isidiokunthii, S. sylvatica, S. phylidiokunthii y S. gallowayana.



CONCLUSIONES
Este trabajo presenta la reconstrucción de la filogenia de los líquenes colombianos pertenecientes al género Sticta tomando como punto de partida el estudio realizado por Moncada et al., (2013). Los resultados de este estudio se pueden explicar en función de las diferencias en el procesamiento de datos, lo que significa que las variaciones identificadas explican los cambios en la constitución y distribución de los clados representados en los árboles filogenéticos que se muestran. Otra diferencia entre el árbol de este trabajo y el realizado en el estudio de Moncada et al. (2013) radica en el uso de un programa computacional de análisis filogenético optimizado y perfeccionado en el tiempo que separa los dos estudios; esto es evidente en los registros de actualización de MegaX (Mega Software, 2022). En estos se enumeran múltiples cambios realizados por la empresa de software para optimizar el funcionamiento del programa, tales como una reconstrucción del algoritmo en febrero de 2018 para correr múltiples iteraciones de forma paralela en la versión 10.0.0 del programa. Múltiples diferencias entre los algoritmos de análisis se pueden hallar en el registro de cambios.1
Como era de esperarse, se observó una separación de las muestras extranjeras (códigos de accesión AY para Estados Unidos) utilizadas como grupo externo, lo que demuestra un buen funcionamiento de los programas computacionales para la construcción de filogenia basándose en información genética. Las especies de origen estadounidense son taxonómicamente más cercanas a U. cavernosa que a las especies del género Sticta de origen colombiano.
Adicionalmente, en el árbol 12 se puede observar la presencia de agrupamientos de manera distinta a lo expuesto en el estudio de referencia antes citado. Se encontraron accesiones que en el árbol de referencia hacen parte de un clado determinado, empero en el árbol consenso del trabajo conforman clados nuevos que tienen ubicaciones diferentes a las que son visibles en el diagrama de Moncada et al., (2013).
Aunque la reconstrucción filogenética que se reporta es sólida desde el punto de vista estadístico, la información lograda no permite construir relaciones entre especies y sus entornos debido a la falta de datos de localización geográfica de las muestras y los vacíos de información complementaria básica de los mismos. Sin embargo, los resultados obtenidos pueden llegar a convertirse en un punto de partida para la construcción de bases de datos que contengan información taxonómica de los líquenes colombianos y que incluya accesiones nuevas que estén acompañadas por registros de datos mucho más completos que puedan ser potencialmente usados para determinar áreas geográficas de distribución estimada. Dejando de lado la carencia de información de georreferenciación, es pertinente afirmar que la reconstrucción del árbol filogenético de las especies Sticta en el presente articulo permitió hallar relaciones taxonómicas nuevas en las accesiones tomadas del articulo de referencia. Esto abre la vía para mayor profundización en investigaciones futuras que incluyan datos de las muestras en su análisis.
1. Link del registro completo de actualizaciones: https://www.megasoftware.net/history
2. Link con los árboles completes https://drive.google.com/drive/folders/1zZp4tg3N7P0SWV18Up0okGOQkkEqW33H?usp=share_link
AGRADECIMIENTOS
Le extendemos un sincero agradecimiento a la profesora Marcela González Ortega y a los estudiantes Santiago González Martínez y Felipe Castellanos Suárez.
REFERENCES
Acevedo Azuero, S., & Charry Castillo, Y. T. (2018). Líquenes como bioindicadores de calidad del aire. Universidad Complutense
Alexopoulos, C., & Mims, C. (1985). Introducción a la Micología. Edit. Omega SA Barcelona, España.
Becedas Mesa, Á. (2021). Análisis computacional de periodicidades en cadenas de ADN nucleosomales.
Cubas, P., Núñez, J., Crespo, A., & Divakar, P. K. (2010). Líquenes: que son y su uso como bioindicadores. GEMM/Proyecto de Innovación, 123, 1-9.
García, L., & Rubiano, L. (1984). Comunidades de líquenes como indicadores de niveles de calidad del aire en Colombia. Cont. Amb, 8, 73-90.
Hawksworth, D. L. (1989). Interacciones hongo-alga en simbiosis liquénicas y liquenoides. In Anales del Jardín Botánico de Madrid (Vol. 46, No. 1, pp. 235-247). Real Jardín Botánico.
Lemey P, Salemi M, Vandamme, A-M. (2009). The Phylogenetic Handbook: A Practical Approach to Phylogenetic Analysis and Hyphotesis Testing. Cambridge University Press.
Mirande, V., & Tracanna, B. C. (2007). Diversidad de cianobacterias, clorofitas y euglenofitas en humedales de altura (Jujuy, Argentina). Lilloa, 39-59.
Moncada, B., Lücking, R., & Suárez, A. (2013). Molecular phylogeny of the genus Sticta (lichenized Ascomycota: Lobariaceae) in Colombia. Fungal Diversity, 64, 205-231.
Moncada, B., Lücking, R., Parnmen, S., & Lumbsch, H. T. (2012). Sticta fuliginosa (Lobariaceae): species or morphotype. In Lichens: From Genome to Ecosystems in a Changing World (The 7th IAL Symposium, Bangkok, Thailand, 9–13 January 2012). Abstract (Vol. 159). DOI 10.1007/s13225-013-0230-0
Suárez, A., & Lücking, R. (2013). Sticta viviana (lichenized Ascomycota: Peltigerales: Lobariaceae), a new species from Colombian paramos. The Lichenologist, 45(2), 153-157.
Spribille, T., Resl, P., Stanton, D. E., & Tagirdzhanova, G. (2022). Evolutionary biology of lichen symbioses. New Phytologist, 234(5), 1566-1582.
Kumar, S., Stecher, G., Peterson, D., & Tamura, K. (2012). MEGA-CC: computing core of molecular evolutionary genetics analysis program for automated and iterative data analysis. Bioinformatics (Oxford, England), 28(20), 2685–2686.
Koichiro Tamura, Glen Stecher, and Sudhir Kumar (2022) Megasoftware: Molecular Evolutionary Genetics Analysis. Update History 38:3022-3027 https://www.megasoftware.net/history